金耳是中国特有的高级滋补养生型食用菌,她以寄生性而在食用菌界特立独行,虽然野生金耳的驯化栽培已有40多年的历史,但是其生物学方面还有许多未解之谜。
图1:金耳子实体
Fig. 1 Fruit-body of Naematelia aurantialba
云南菌视界生物科技有限公司是一家致力于将中国特有物种金耳的野生种质资源进行产业化开发的专业公司,公司研发中心在应用营养生理学、N15标记和栽培学的方法验证了藏穆先生在发表金耳新种时就定义金耳是寄生型真菌的正确结论的基础上,近日以两篇网络首发论文的形式揭示了金耳(菌)有性生殖的密码,丰富了金耳的生物学基础知识,为金耳的遗传育种研究奠定了坚实的基础,将大大促进金耳的产业化发展步伐。
以曹瑶为第一作者,李荣春为通讯作者,9月7日发表在《食用菌学报》的“采用单孢杂交技术判定金耳极性”论文用酵母状单孢分离物,经过四轮交配实验确定金耳单孢分离物具有4种交配型(A1B1、A1B2、A2B1和A2B2),表明金耳属于四极性异宗结合真菌。用交配及栽培实验,验证了只有当2个单孢分离物的A、B交配型均不同时交配形成的双核体方可生长发育形成金耳子实体。
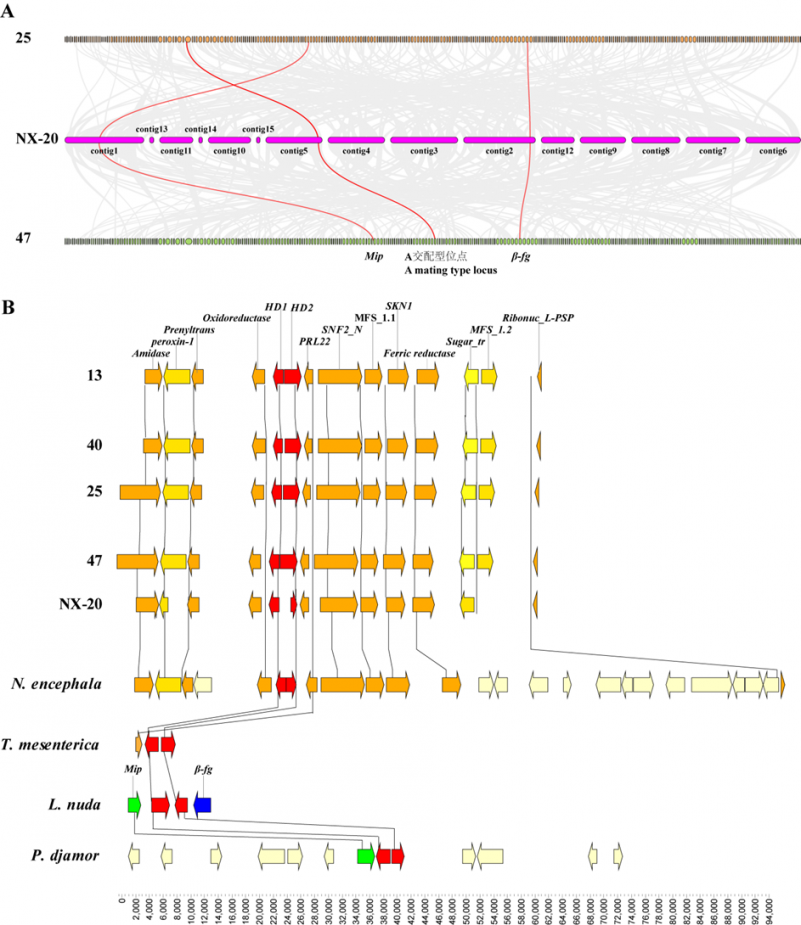
图2:金耳基因组共线性分析(A);不同真菌的A交配型位点共线性分析(B)Fig. 2 Synteny analysis of Naematelia aurantialba genome (A); Synteny of A mating locus of different fungus (B).

以沈真辉和曹瑶为共同第一作者,李荣春为通讯作者,10月25日发表在《菌物学报》的“基于基因组数据解析金耳A和B交配型位点结构”论文对前一篇论文研究获得的4种不同交配类型的金耳单核菌株,利用二代测序技术对基因组进行重测序,通过生物信息学方法找到了金耳的A和B交配型位点,并与其它大型真菌进行比较。结果显示4个单核体中含有2种A交配型位点和2种B交配型位点;其中2种A交配型位点都包含一对HD1和HD2基因,以典型的“头对头”方式排列,其上下游具有保守的氧化还原基因和PRL22基因,而mip和β-fg基因不位于HD基因两侧,不存在紧密连锁关系。2种B交配型位点均含有1个信息素受体(STE3)和1个信息素前体基因(phB),比较B1和B2位点发现,B1位点的STE3和phB基因紧密排列,在phB上游紧密相邻着1个B2位点没有的RVT_1基因, 在B2位点中STE3和phB基因之间插入了3个基因,其中STE12基因在B1位点没有,与其他真菌相比,B位点共线性较差,表明不同真菌中B位点变异性较大;在4个单核体中均含有信息素受体NaSTE3-3基因,但在其上下游未发现信息素前体基因,并与B交配型位点不在同一contig上。
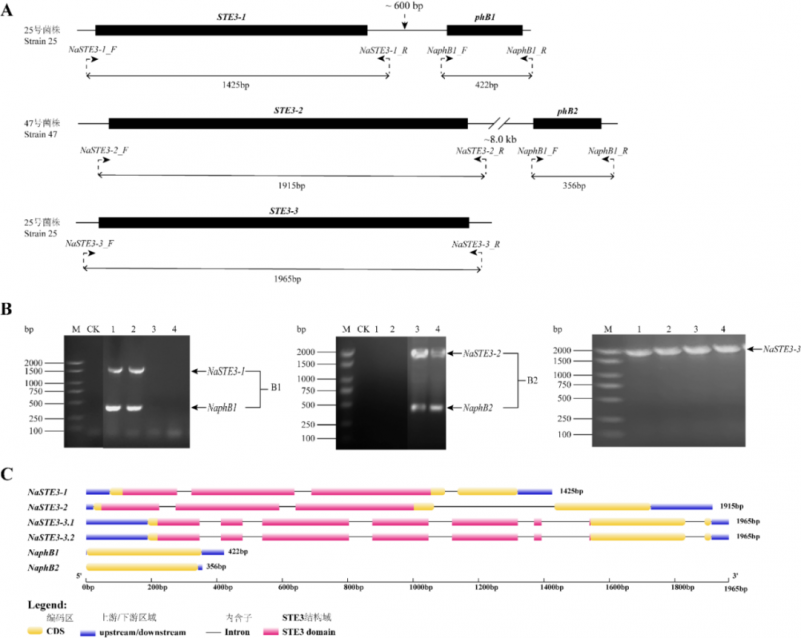
图3: 金耳B交配型位点和NaSTE3-3基因所设计的引物及扩增片段的相对位置(A);PCR扩增图谱(B)和基因结构图(C) M:DL2000 Marker;CK:水对照;泳道1-2:13和25号单核体菌株;泳道3-4:40和47号单核体菌株
Fig. 3 Relative positions of designed PCR primers and amplified DNA fragments (A); The PCR amplification validation (B) and gene structure map (C) of B mating type locus structure and NaSTE3-3 of Naematelia.aurantialba. M: DL2000 Marker; CK: Water control; lanes 1-2: Monokaryotic strains 13 and 25; lanes 3-4: Monokaryotic strains 40 and 47.
两篇论文全面系统地揭示了金耳菌生活史中有性生殖的关键密码,阐述清楚了金耳菌的交配系统类型及特征,全面解析了金耳菌的A、B交配基因的结构,在两篇论文发表的同时,相关成果还获得了两项授权的发明专利,为金耳生活史、遗传和育种等研究奠定了科学基础。两篇论文的发表,为公司更好地理解和认识金耳,为公司3.0版的金耳栽培技术的开发,为公司致力于将“云南野生金耳资源转化成为老百姓餐桌上的健康食品、美味佳肴”的梦想注入了知识源泉。下一步,公司将对金耳的基因组、转录组和代谢组进行深入研究,对金耳野生种质资源进行全面系统的评价,开展金耳的传统选育、太空诱变育种、杂交育种、基因组育种以及基因编辑育种等新品种育种研究,不断揭示更多的金耳密码。(来源:菌视界)
 APP下载
APP下载